
Ethical issues and permits
Permits to implement the experiment were granted from the French Ministry of Research (APAFIS #45548-2023102712468223) after evaluation by the Comité Régional d’Ethique d’Occitanie and from Préfecture des TAAF (A-2024-95) after evaluation by the Conseil de l’Environnement Polaire and Conseil National de la Protection de la Nature. After the suspicion of HPAI emergence on Crozet, permits were granted by the Préfecture des TAAF to pursue eco-epidemiological fieldwork (A-2024-144).
Model species and study population
The king penguin is a long-lived colonial seabird, breeding in colonies that can reach tens to hundreds of thousands of individuals, with an extended reproductive cycle of ca .14 months35,60. A single egg is laid, followed by an incubation time of 54 days and an extended rearing period of c.11 months. Partners share alternatively the incubation of the egg, then the care of the chick, until its full thermal emancipation (around 30–40 days post-hatching). Chicks are then left ashore in the colony by their parents for several months before fledging to sea at around 300 days post-hatching. Once the chicks are emancipated, they often gather in groups (‘crèches’) to withstand the harsh conditions of the Austral winter. Over that time period, they can be captured, resighted and recaptured, allowing good conditions for their monitoring over several months61.
The experimental vaccination trial on king penguin chicks was conducted from February to November 2024 on Possession Island (Crozet archipelago, 46° 24′ 41″ S, 51° 45′ 22″ E, Southern Indian Ocean), within the colony of Baie du Marin.
saRNA vaccine and practical issues
The saRNA RESPONS AI H5 vaccine (Ceva Santé Animale) was used to explore the immune response of the king penguin chick against a H5 HPAI clade 2.3.4.4b protein in natural settings. The vaccine was provided by the producer under a Material Transfer Agreement (MTA) for exclusive use in the trial. Stock vaccine vials were transported and stored at −80 °C during the whole travel to reach the lab facilities on Possession Island, until processing for the field experiment.
Prior to any injection, stock vaccine vials were placed at 4 °C for 12 h before each session of injections, to ensure a complete defrosting. The thawed stock vaccine solution was then diluted freshly with a provided diluent according to the manufacturer’s guidelines. The diluted vaccine was administered within the recommended 2 h post-dilution (0.2 ml intra-muscular injection of 3 µg in the thigh). The use of the saRNA vaccine under field conditions in sub-Antarctica was facilitated by the proximity of the study colony to the scientific station, where −80 °C storage facilities were available. However, the possibility to keep the stock vaccine vials at 4 °C during 24 h before conducting a session of injections could allow some flexibility to organise vaccination trials in remote colonies.
Experimental vaccination design
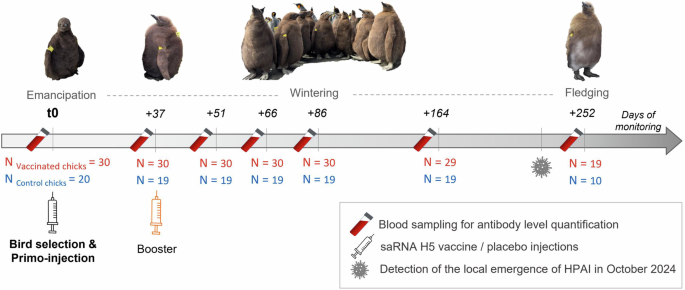
For the experiment, 50 emancipated chicks of approximately 45 days post-hatching were selected and randomly assigned to two treatment groups, and they were monitored along their rearing period, until fledging to sea. Those chicks were temporally captured and tagged in batches using alphanumeric plastic flipper bands and coloured flags allowing efficient and individual tracking during the whole chick rearing-period. Microtransponders (BioTherm13® 13 mm 134.2 kHz ISO FDX-B, Biomark, Idaho, USA) were subcutaneously inserted into the back of the neck, each one related to a unique and permanent identification number. Birds received either a primo-injection (t0) of the saRNA H5 vaccine in the thigh muscle (n = 30 chicks, vaccinated group), or a placebo injection using the vaccine diluent (n = 20 chicks, control group). A boost-injection was applied approximately 37 days later to all vaccinated chicks, while control chicks received a second placebo injection, under the same handling conditions as at t0. The bird immune responses were monitored along the entire rearing period through blood sampling at 7 capture times. For clarity of presentation and statistical analyses, sampling days were grouped into seven categories: t0, t0 + 37 days, t0 + 51 days, t0 + 66 days, t0 + 86 days, t0 + 164 days and t0 + 252 days (Fig. 1; and Supplementary Table 12). Blood samples (0.2 to 1 ml) were collected from the ulnar vein using a 1 ml heparinized syringe. Plasmas were separated from red blood cells by centrifugation within a few h. They were immediately frozen and stored at −20 °C before conducting serological analyses. All chick external tags (coloured flags and flipper bands) were removed at the end of the experiment, at t0 + 252 days. Both groups of control and vaccinated penguin chicks likely contained animals of both sexes, but immune responses of the animals were not differentiated by sex. The immune response of the mixed-sex animal group as a whole was analysed.
Evaluation of the vaccine safety
To assess potential adverse effects of the vaccine, we first evaluated local short-term effects after each intra-muscular injection: chicks were kept a few minutes before release to check for any sign of limping, local swelling, or typical distress behaviour (hyperventilation, lack of reactivity…). Chick condition was also monitored at each capture. The chick survival rate in both treatment groups was evaluated regularly along the entire trial through remote checks from the edge of the colony, until all chick external tags were removed at t0 + 252 days. We assessed potential long-term adverse effects by monitoring the change in chick body mass and body size (flipper length), as morphometric indicators of chick growth, until t0 + 164 days. In the context of the HPAI outbreak detected locally in late October 2024 on Possession Island, we could not record the chick body masses/sizes at the last sampling point (t0 + 252 days) due to biosecurity restrictions. To evaluate whether vaccination affected chick body growth trajectories, we used a Bayesian linear mixed-effects model implemented in the brms package (version 1.22.0) of the R software49. The model included body mass or flipper length as the response variables, with days post-primo-injection, treatment group (vaccinated vs. control), and their interaction as fixed effects. Chick identity was included as a random intercept to account for repeated measures of individuals. To formally assess the absence of a biologically meaningful difference between treatment groups over time, we used a Region of Practical Equivalence (ROPE) approach. The ROPE for the interaction term (days: treatment) was set to ±0.2 kg. Posterior distributions and ROPE inclusion percentages were computed using the bayestestR package (version 0.16.0).
To evaluate whether the vaccination treatment had affected the chick survival over time, we fitted a Cox proportional hazards model with treatment group (vaccinated vs. control), as a categorical predictor. Time-to-event was measured from t0 to the last capture at t0 + 252 days, when external identification of individuals where removed. We used the coxph() function from the survival package of the R software (version 3.8-3). We assessed the proportional hazards assumption using Schoenfeld residuals. Model results are reported as hazard ratio (HR) with 95% confidence interval (CI). A two-sided p-value < 0.05 was considered statistically significant.
Quantification of specific antibody levels
The production and temporal persistence of specific H5 antibody levels were measured in plasma samples using commercially-available enzyme-linked immunosorbent assays (ELISA) kits. First, plasma samples were analysed using a competitive ELISA assay targetting the H5 protein of the clade 2.3.4.4b virus (H5-cELISA) and suitable for the monitoring of vaccination of domestic and wild birds (ID Screen® Influenza H5 Antibody Competition 3.0 Multi-species, reference no. FLUH5V3-5P, Innovative Diagnostics SARL, Grabels, France). Given previous results obtained on other seabird species suggesting that anti-chicken conjugate antibodies could bind with seabird antibodies62, we complemented our results using an anti-H5 indirect ELISA assay (H5-iELISA) (ID Screen® Influenza H5 Indirect, reference no. FLUH5S-2P, Innovative Diagnostics SARL, Grabels, France). In order to evaluate the natural exposure of chicks to unspecific influenza A viruses potentially circulating in the colony (DIVA approach), we also tested plasma samples using a competitive ELISA assay targetting the non-specific nucleoprotein (NP-cELISA) (ID Screen® Influenza A Antibody Competition Multi-species, reference no. FLUACA-5P, Innovative Diagnostics SARL, Grabels, France). ELISA assays were conducted according to instructions provided by the manufacturer. Plasma anti-H5 and anti-NP antibody levels were obtained by reading absorbance values at 450 nm using a microplate reader (Tecan Infinite® 200 Pro; Tecan Group Ltd., Mannendorf, Switzerland), and correcting raw absorbance values using optical densities (OD) of positive (PC) and negative (NC) controls provided for each kit (ODPC and ODNC, respectively). Anti-H5 and anti-NP antibody levels obtained with competitive ELISA are expressed as inhibition values (inhibitionsample = ODsample/ODNC), while anti-H5 antibody levels obtained with indirect ELISA are expressed as S/P ratio (S/Psample = ODsample/ODPC). We referred to each threshold value provided by each ELISA kit instructions to discriminate seropositive from seronegative samples: H5-cELISA = 0.50; H5-iELISA = 0.50; NP-cELISA = 0.45. Analyses conducted on repeated sub-samples allowed us to assess the between- and within-assay repeatability for each ELISA test (see Supplementary Table 13 for detailed coefficients of variation).
HP H5N1 seroneutralisation assay
A seroneutralisation assays was ran on a subset of plasma samples to explore the capacity of specific antibodies to seroneutralise a HPAI H5 virus of clade 2.3.4.4b. The assay was conducted under Biosafety Level 3 conditions using the A/chicken/France/D2107428/2021_2.1b H5N1 HP virus from 2.3.4.4b clade. Plasmas were heat-inactivated (56°C for 60 min) and twofold serial dilutions were performed in a 50-μl volume in microtiter plates. Sera obtained from experimentally immunized chicken with the homologous inactivated (Sp_H5N1_A/chicken/France/D2107428/2021_1) (positive control serum) and obtained from SPF chicken (negative control serum) were also used. Diluted plasmas were mixed with an equal volume containing 100 TCID50 of the H5N1 virus. Four control wells of virus alone (positive for the cytopathogenic effect (CPE) control) and four control wells of dilution medium alone (negative CPE control) were included. After 1h incubation at 37°C and 5% CO2 for neutralisation, 100 μl of MDCK cells at 2 × 105/ml were added to each well. Plates were incubated at 37°C and 5% CO2 for 72h, after which the CPE was read under the microscope. For each plasma, the seroneutralisation titer was expressed as the mean of the last dilution factor for which no CPE was observed of two independent experiments. The positive control serum has a titer greater than 2048, the negative control serum has a titer less than 8.
Antibody levels comparison
Statistical analyses were conducted using R63 (R Core Team, 2023, version 4.3.1). We compared the antibody level at each time point between vaccinated and control individuals using analyses of variance. The same model was used for both competition and indirect ELISA results. To assess the effect of vaccination on antibody dynamics, we used linear mixed-effects models (LMMs) fitted with the lme4 package in R. Antibody levels were modeled as the response variable, with treatment group (vaccinated vs. control), sampling day (as a factor), and their interaction as fixed effects. Individual identity was included as a random effect to account for repeated measures.
We tested the significance of main effects and interactions using Type III ANOVA with Satterthwaite’s approximation for denominator degrees of freedom, implemented in the lmerTest package. When significant interactions were detected, we conducted post-hoc pairwise comparisons between treatment groups at each sampling day using estimated marginal means (EMMs, emmeans package), with Tukey adjustment for multiple testing.
Fitted models for antibody levels
To model the dynamics of anti-H5 antibody levels, we fitted the sequential values from each individual with a Holling IV rational function64 in order to account for the expected sigmoid early build-up of antibody level following the primo- and boost-injections, followed by a decay of antibody level towards a potential asymptotic value for large numbers of days post vaccination. A fixed treatment effect was set to compare the vaccinated and control groups (stable low level was expected for control chicks).
We fitted a non-linear regression model to the time series data (exact time series) using the nlsLM() function from the minpack.lm R package (version 1.2-4). The Holling IV function adapted to our modelling situation is:
$$f\left({\mbox{days}}\right)=\frac{{\mbox{a}}\cdot {\mbox{day}}{{\mbox{s}}}^{2}}{{\mbox{b}}+{\mbox{c}}\cdot {\mbox{days}}+{\mbox{day}}{{\mbox{s}}}^{2}}$$
(2)
with f(days), the antibody level (calculated from ELISA titre values); days, the number of days since the primo injection; a, the value of the function at the asymptotic reached at high values of days; (−2b/c), the value of day at the peak antibody level. We compared a pooled model, in which vaccinated and control groups shared the same temporal response function, to a group model in which curve parameters were allowed to vary between the two groups (vaccinated vs placebo).
We compared three competing models:
-
Pooled model: vaccinated and control chicks shared the same response curve (no group effect);
-
Full group model: curve parameters were allowed to differ between groups, with the following parameterization:
$$f({days})=\frac{({{\rm{a}}}+{{\rm{da}}}\cdot {{\rm{treatment}}})\cdot {{{\rm{days}}}}^{2}}{(b+{db}\cdot {treatment})+(c+{dc}\cdot {treatment})\cdot {days}+{{{\rm{days}}}}^{2}}$$
(3)
where days is the number of days since the primo-injection and treatment is a binary indicator of group membership (0 = control, 1 = vaccinated). We parameterized the Holling type IV function such that the control group served as the reference, with parameters acontrol, bcontrol and ccontrol. For the vaccinated group, parameters were expressed as additive deviations from the control: acontrol+da, bcontrol+db, and ccontrol+dc.
Model comparisons were conducted using the corrected Akaike Information Criterion (AICc) to account for differences in model complexity and sample size, using the AICcmodavg R package (version 2.3-4). The model with the lowest AICc value was retained as the most parsimonious representation of the data. A difference in AICc greater than 2 was considered evidence of meaningful support for the better-fitting model.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.