
In this study we showed that standard in vitro culture of primary mouse microglia precipitates a rapid and profound loss of homeostatic features within 24 h of plastic adherence, as evidenced by downregulation of signature transcripts (Tmem119, P2ry12, Cx3cr1, Hexb, Fcrl2, Olfml3, Tgfbr1). Critically, this “culture shock” can be partially reversed by reconstituting key CNS niche signals – hTGF-β1, hIL-34, and hCX3CL1 – together with collagen IV coating, cholesterol and insulin-transferrin-selenium in serum-free conditions, yielding both transcriptional recovery (particularly Tmem119, Olfml3, Cx3cr1, Tgfbr1) and ramified morphology consistent with a more resting-like microglial state. These results address the translational gap outlined in the introduction by demonstrating that microenvironmental inputs essential for microglial homeostatic features in vivo can be leveraged to partially preserve these features in primary microglia ex vivo6,7,27. We used neonatal primary microglia and therefore confine our conclusions to neonatal-derived cultures. Although developmental stage shapes baseline transcription, the direction of cytokine and ECM responses aligns with reports in adult microglia4,8.
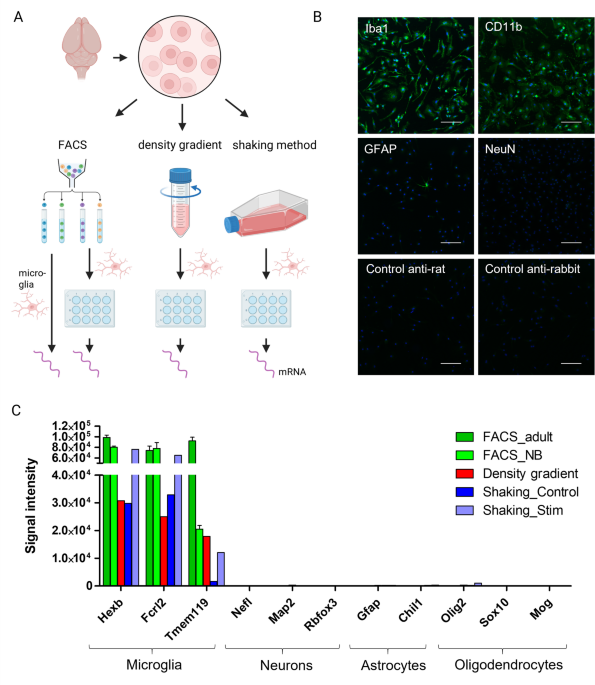
An important control in our study was the comparison across isolation methods – FACS, Percoll, and shaking – which all converged on the same rapid loss of homeostatic transcripts after 24 h, whereas freshly FACS-sorted adult microglia maintained high expression of signature genes at baseline. This points to culture context as the dominant driver of transcriptional drift, rather than the isolation workflow per se, consistent with environment-dependent microglial identity7. Over a 7-day time course, we observed further progressive reductions for a subset of genes (Cx3cr1, Hexb, P2ry12), suggesting a continuing adaptation to the artificial culture milieu and highlighting the need for sustained niche support to prevent phenotypic erosion7,8.
Because the ultimate goal was to approximate adult microglial physiology in vitro, particularly in the context of neuroinflammation and neurodegeneration, we used freshly isolated adult microglia as the reference baseline for our RT-qPCR analyses. Neonatal microglia are technically more accessible and yield higher cell numbers, but they differ from adult microglia in their baseline transcriptional, morphological, and functional properties. We therefore included neonatal ex vivo microglia as a separate reference at the transcriptome level (Fig. 4A,B) and interpret our data as assessing how far neonatal-derived cultures can be driven toward adult-like homeostatic transcriptional signatures, rather than implying equivalence between neonatal and adult microglia.
Our observations are congruent with prior work showing that microglial phenotype is tightly governed by CNS-derived cues and is rapidly reprogrammed upon removal from their tissue context4,8. Immediate ex vivo profiling revealed that microglia possess a distinct homeostatic transcriptional network in vivo which collapses in conventional culture, with downregulation of Tmem119 and P2ry12 and induction of activation and proliferative programs7,27. Serum-containing media are a major driver of this drift, promoting amoeboid morphology, proliferation, heightened phagocytosis, and broad activation signatures8,28. In line with these reports, we found that serum exposure favored amoeboid microglia, whereas microglia cultured under serum-free conditions yielded thin, highly branched processes typical for surveillant microglia. However, serum withdrawal alone reduced signature gene expression, underscoring that phenotype stabilization requires both removal of activating stimuli and active provision of pro-homeostatic signals. We used insulin-transferrin-selenium (ITS) to partially replace serum by providing defined metabolic support and maintaining baseline viability. Selenium, supplied as part of ITS, is essential for antioxidant defense in microglia, especially through its role in glutathione peroxidase 4 (GPX4), protecting cells from oxidative stress and ferroptosis29,30. Insulin regulates microglial glucose uptake and homeostatic metabolism primarily via IRS-1/PI3K/AKT signaling pathways, which are crucial for maintaining microglial quiescence and function31,32. Transferrin ensures sufficient iron availability for cell proliferation and enzymatic reactions; limiting free iron is also neuroprotective33,34. We did not directly quantify proliferation or phagocytic activity in our cultures, so our conclusions are limited to transcriptional and morphological endpoints.
Building on prior work that used CNS niche cytokines to stabilize primary microglia4,8, our data indicate that hTGF-β1 is the predominant driver of transcriptional rescue, while hIL-34 and hCX3CL1 were included to represent neuronal survival and restraint signals. The three-factor mix improved morphology and shifted global profiles toward neonatal ex vivo microglia, without implying additive or dominant effects of hCX3CL1 or hIL-34. Specifically, TGF-β signaling is indispensable for microglial development and maintenance of the homeostatic signature (including Tmem119 and P2ry12) in vivo and can restore aspects of microglial identity ex vivo19. The canonical Smad-dependent TGF-β pathway acts as a central regulator of microglial maturation, ensuring their differentiation from progenitors and sustaining a quiescent, surveillant phenotype under physiological conditions19. Genetic ablation of TGF-β signaling in microglia leads to a loss of homeostatic markers and induces a transcriptomic shift toward a disease-associated or primed state, highlighting its role in preventing maladaptive activation19. IL-34, a high-affinity ligand for CSF1R, is a key survival factor for microglia, and sustained CSF1R signaling is necessary for microglial viability and appropriate transcriptional programs35,36. The neuronal chemokine CX3CL1 (fractalkine) restrains microglial activation via CX3CR1 and modulates microglia–synapse interactions, thereby supporting surveillance and limiting inflammatory responses37,38. Furthermore, protocols for human iPSC-derived microglia typically combine IL-34 and TGF-β to recapitulate core aspects of microglial identity, supporting the translational relevance of our cytokine cocktail approach10,39,40. Together, these literature data provide a biological rationale for the restoration of signature transcripts we observed when hTGF-β1, hIL-34, and hCX3CL1 were supplied.
Beyond these soluble cues, additional contributions from extracellular matrix shaped the magnitude of rescue: we observed that coating of cell culture wells with 2 µg/mL collagen IV augmented the effects of the cytokine cocktail on selected microglial genes and preserved the ramified morphology. Collagen IV is a principal component of vascular basement membranes, that microglia engage via integrin-dependent focal adhesions, thereby modulating their adhesion, mechanosensation, and directed motility at the blood-brain barrier41. Accordingly, presenting collagen IV in vitro may help sustain the adhesion, survival, and homeostatic phenotype of primary microglia by mimicking the basement membrane environment of the neurovascular unit and providing integrin-mediated signaling cues that support ramified morphology and anti-inflammatory functions41. Besides that, cholesterol availability is a key determinant of microglial identity: microglia sense neuronal cholesterol via Trem2, a receptor essential for microglial metabolic fitness, which regulates ApoE-dependent lipid processing42. However, local excessive amounts of cholesterol in the CNS, for example during the breakdown of myelin, can lead to cholesterol transport capacity being overwhelmed. This can induce stress responses in microglia and cause a disease-associated transcriptional state43. In this study, 1.5 µg/mL cholesterol did not increase the majority of homeostatic genes examined and affected Tgfbr1 only in the presence of cytokines and collagen IV. We did not measure survival directly, and inclusion was based on prior reports of improved viability8.
Experimental approaches exist in which primary microglia are cultivated alongside astrocytes, neurons, or both44,45,46. Factors secreted by these cells into the culture medium support the ramification of the microglia and induce the expression of genes that are typically lost in vitro. The disadvantage of these approaches is that they are costly and time-consuming to set up, as up to three different cell types must be provided. By contrast, our protocol enables the easy and rapid addition of selected CNS niche cues, offering a simplified solution that can produce similar beneficial effects on cultured microglia morphology and homeostatic gene expression, although we did not directly compare our conditions to such co-culture systems.
Several conceptually related studies have previously combined CSF1R ligands, TGF-β and cholesterol in serum-free conditions to support rodent8 or human47,48 microglia. Our work builds on these protocols but extends them in three main ways. First, we systematically quantified a defined set of homeostatic genes by RT-qPCR across multiple isolation methods and culture conditions, demonstrating that the same rapid culture-induced loss and partial rescue are observed regardless of the initial isolation workflow. Second, we validated the impact of the optimized condition transcriptome-wide using microarray and RNA-seq, showing that most genes from the Butovsky homeostatic gene set are upregulated and that inflammatory and stress-associated programs are attenuated relative to serum-free control medium. Third, we provide a practical, collagen IV–based, chemically defined, serum-free recipe that can be implemented without co-culture and is specifically optimized for neonatal primary microglia. In particular, whereas Bohlen et al. primarily characterized microglial survival, morphology and phagocytic capacity under defined serum-free conditions, our data provide the complementary genome-wide transcriptional landscape of microglia exposed to conceptually similar CNS niche cues, thereby linking functional and molecular readouts and offering an experimentally accessible workflow for stabilizing homeostatic-like signatures in primary culture.
Several limitations should be considered when interpreting our findings. First, microglial enrichment was assessed by immunocytochemistry for shaking-derived cultures at early time points and by transcriptomic analysis of lineage marker expression in selected conditions, which indicated very low expression of neuronal, astrocytic and oligodendrocytic markers relative to microglial signature genes. However, we did not systematically quantify contamination across all isolation methods, conditions and time points, so minor contributions from other brain-resident cell types cannot be completely excluded. Second, our ex vivo reference transcriptomes were obtained after enzymatic digestion in the presence of FBS and without transcriptional inhibitors such as actinomycin D, and therefore likely already reflect some isolation-induced changes; they should be interpreted as approximations to the in vivo state rather than undisturbed in vivo profiles. Third, our primary readouts were mRNA expression and morphology. We did not directly assess functional endpoints such as phagocytic capacity, cytokine secretion or proliferation, and therefore do not claim that these aspects of microglial function are fully preserved under our optimized conditions. Future studies should complement our transcriptomic and morphological analyses with functional assays (e.g., phagocytosis assays, cytokine secretion measurements, and Ca²⁺-signaling or calcium-imaging readouts) to determine to what extent the optimized serum-free, collagen IV–based conditions also preserve microglial functions relevant to neuroinflammation and CNS homeostasis. Fourth, RT-qPCR normalization relied on single reference genes (Gapdh or Ywhaz), whose expression can be modulated by activation state and culture condition; although the major expression trends observed by RT-qPCR were consistent with those from microarray and RNA-seq, the microarray and RNA-seq datasets were normalized using global approaches that do not rely on individual housekeeping genes and therefore provide an independent reference for the direction of expression changes. The use of single housekeeping genes in our RT‑qPCR assays nevertheless represents a methodological limitation that should be addressed in future work by validating multiple reference genes.
In summary, we provide evidence that the rapid loss of microglial homeostatic features after plastic adherence is not irreversible: a defined set of CNS niche cues – hTGF-β1, hIL-34, and hCX3CL1 – combined with ECM and lipid support, can partially restore homeostatic-like transcriptional signatures and ramified morphology in cultured neonatal primary microglia. This optimized, scalable workflow is expected to improve physiological relevance in vitro compared with conventional serum-containing culture, mitigate culture-induced transcriptional artifacts, and enable more robust interrogation of microglial mechanisms implicated in neuroinflammation and synapse regulation across disease models.